NEJM
Review
|
Cornelia M. Weyand, M.D., Ph.D., and J�rg J. Goronzy, M.D., Ph.D.
Inflammation of large arteries such as the aorta and its major branches occurs in a number of disorders, including Kawasaki's syndrome, Beh�et's syndrome, rheumatoid arthritis, syphilis, and tuberculosis. However, aortitis and large-vessel arteritis are characteristics of two entities � giant-cell (temporal) arteritis and Takayasu's arteritis. These arteritides involve similar histologic abnormalities1 but differ in the age of onset and the vascular structures that are preferentially targeted.2,3 They share pathogenic pathways that distinguish them from other vasculitides. Cellular immune responses involving T cells, antigen-presenting cells, and macrophages are fundamental elements in giant-cell arteritis and Takayasu's arteritis, and there is no evidence of an autoantibody component.
Giant-cell arteritis affects almost exclusively persons older than 50 years of age, and disease risk is highest among those who are 75 to 85 years of age.4 Disease susceptibility has been associated with northern European descent, and two thirds of those affected are women. Incidence rates reach 15 to 25 cases per 100,000 persons over the age of 50 years.4,5 Giant-cell arteritis typically causes vasculitis of the extracranial branches of the aorta and spares intracranial vessels. Transmural inflammation of the arteries induces luminal occlusion through intimal hyperplasia. Clinical symptoms reflect end-organ ischemia.3,6,7 Branches of the external and internal carotid arteries are particularly susceptible. Their involvement leads to the classic manifestations of blindness, headache, scalp tenderness, and jaw claudication. Vasculitis of the vertebral arteries can impair the posterior cerebral circulation and cause stroke, transient ischemic attacks, vertigo, and dizziness. Involvement of the subclavian, axillary, and proximal brachial arteries leads to the aortic arch syndrome of claudication of the arms and absent or asymmetric pulses.8 Whereas giant-cell arteritis of medium-sized arteries results exclusively in narrowing and obstruction of vessels, the clinical complications of aortitis, which most often occurs in the thoracic aorta, are aneurysm formation, dissection, hemorrhage, and rupture.9,10
In almost all patients with giant-cell arteritis, a syndrome of systemic inflammation accompanies vascular manifestations. The nonspecific symptoms include malaise, anorexia, weight loss, fever, night sweats, and depression. In patients with highly elevated acute-phase responses, such as a high erythrocyte sedimentation rate and increased levels of C-reactive protein, these symptoms are important clues to the diagnosis. Signs of systemic inflammation are also present in patients with polymyalgia rheumatica, a disorder characterized by severe myalgias and stiffness of the muscles of the neck, shoulder girdle, and pelvic girdle. About 30 to 40 percent of patients with giant-cell arteritis also have polymyalgia rheumatica. The two conditions affect the same population of patients, have the same genetic risk factors,11 and have similar patterns of acute-phase responses.12
Takayasu's arteritis, also known as pulseless disease, primarily involves large elastic arteries. Adolescent girls and women in their second and third decades of life are at highest risk.2 The syndrome is most commonly seen in Japan, Southeast Asia, India, and Mexico.2 In North America, the annual incidence is estimated to be 2.6 per million people.13 Panarteritic inflammatory infiltrates cause marked thickening of the involved artery and subsequent luminal narrowing and occlusion. Diffuse dilatation, formation of aneurysms, and thrombosis occur more frequently in Takayasu's arteritis than in giant-cell arteritis. Clinical manifestations range from asymptomatic disease with impalpable pulses to catastrophic neurologic impairment.14,15 Most patients have diminished pulses or no pulses in the arm and blood-pressure discrepancies between the right and left arms owing to subclavian stenosis. Involvement of the carotid arteries leads to cerebral ischemia and is manifested clinically as stroke, postural dizziness, seizures, and amaurosis. Aortic regurgitation results from dilatation of the ascending aorta. The presence of hypertension most often reflects renal-artery stenosis. The long-term prognosis is frequently influenced by the presence of congestive cardiac failure associated with hypertension, aortic regurgitation, or dilated cardiomyopathy. Cough, dyspnea, chest pain, and pulmonary hypertension are present in patients with pulmonary involvement. Stenotic lesions at the origin of the renal and mesenteric arteries can complicate cases involving the abdominal aorta. As in giant-cell arteritis, vascular disease in Takayasu's arteritis is accompanied by fever, malaise, arthralgias, myalgias, weight loss, and anemia.
Despite differences in presentation and the clinical course, giant-cell arteritis, polymyalgia rheumatica, and Takayasu's arteritis share many pathogenic principles, and similar rules apply in the diagnostic and therapeutic approaches. The following discussion of pathogenic pathways will focus mostly on giant-cell arteritis, a reflection of the accessibility of inflamed vascular tissue that has facilitated mechanistic studies.
The Pathogenic Model of Giant-Cell Arteritis
The diagnosis of giant-cell arteritis is established by biopsy, usually of the temporal artery. The findings are those of a panarteritis with mononuclear infiltrates penetrating all layers of the arterial wall (Figure 1A). Typically, activated T cells and macrophages are arranged in granulomas. Multinucleated giant cells, when present, are usually close to the fragmented internal elastic lumina (Figure 1B). Often, the intimal layer is hyperplastic, leading to concentric occlusion of the macrolumen.
|
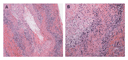
Progress in understanding giant-cell arteritis has derived from three key observations. First, it is now clear that giant-cell arteritis is a T-cell�dependent disease.16,17 The CD4+ T cells that orchestrate the injury of tissues are a sine qua non in the vasculitic process. Second, T-cell activation in the nonlymphoid environment of the arterial wall requires the activation of specialized antigen-presenting cells, the dendritic cells.18 The activation of monocytes and macrophages is responsible for the systemic inflammatory syndrome in giant-cell arteritis and polymyalgia rheumatica.19 Third, the blood vessel determines the site specificity of giant-cell arteritis. Resident cells of the arterial wall respond uniquely to the immune injury mediated by tissue-infiltrating cells.20 The end result is an occlusive vasculopathy caused by the rapid proliferation of the intima or the formation of an aortic aneurysm caused by destruction of the arterial wall.
T-Cell�Dependent Vasculitic Inflammation in Giant-Cell Arteritis
The CD4+ T cell is the critical cellular player in the vasculitic lesion.21,22,23 Direct evidence of a central role of T cells comes from experiments in a mouse chimera model.22 These chimeras are created by engrafting temporal-artery�biopsy specimens from patients with giant-cell arteritis into mice with severe combined immunodeficiency. Antibody-mediated depletion of tissue-infiltrating T cells in such chimeras abrogates essentially all inflammation in the implanted human tissue.22 Certain allelic variants of class II antigen-presenting HLA molecules constitute a genetic risk factor for giant-cell arteritis and polymyalgia rheumatica,24 providing indirect evidence of the crucial role of antigen stimulation of CD4+ T cells.11 Moreover, clonally expanded populations of CD4+ T cells with identical antigen receptors have been isolated from distinct vascular lesions of the same patient.25 Such clonal expansion is characteristic of antigen-induced proliferation.
The antigens recognized by CD4+ T cells in giant-cell arteritis are unknown. T cells isolated from vascular infiltrates proliferate in response to extracts of temporal arteries from patients with giant-cell arteritis or polymyalgia rheumatica,26 suggesting the existence of a shared antigen. Infectious agents, toxins, drugs, and autoantigens in arteries would qualify, and parvovirus, influenzavirus, varicellavirus, and Chlamydia pneumoniae have been suspected.27,28,29,30 Reports of material of infectious origin in temporal arteries have, however, not been corroborated.31,32
The concept that giant-cell arteritis is the consequence of antigen-specific T-cell responses in arterial tissue implies three critical events: T cells gain access to a site that they usually do not enter, an inciting antigen is accessible, and antigen-presenting cells that are capable of T-cell stimulation differentiate.
T cells leave the blood and enter tissue only if endothelial cells are activated and chemokines are produced in the tissue. In giant-cell arteritis, the port of entry is the vasa vasorum in the adventitia and not the macroendothelium. Microvessels in the inflamed arteries produce a variety of adhesion molecules that regulate leukocyte transport.33,34,35 Activated dendritic cells produce the chemokines CCL19 and CCL21, which have a critical role in attracting T cells and macrophages into the arterial wall.18 It seems that activation of endothelial cells and dendritic cells precedes T-cell recruitment to the tissue and that the activation and differentiation of dendritic cells are key events in the pathogenesis of giant-cell arteritis.18
Vessel-Wall Inflammation
Dendritic cells have only recently been recognized as a resident population in healthy medium-sized arteries, where they are located exclusively in the adventitia.18 Adventitial dendritic cells are positioned at the medial border, often outside the external elastic lamina. In healthy arteries, dendritic cells have a gene-expression profile characteristic of that of an immature cell (Figure 2A). Immature dendritic cells have a role in maintaining T-cell unresponsiveness. In contrast to mature dendritic cells that induce T-cell responses, immature dendritic cells do not express the costimulatory molecules CD80 and CD86. If T cells encounter antigen on immature dendritic cells, they receive an inhibitory (tolerizing) signal.36,37,38,39 The function of adventitial dendritic cells may therefore lie in preventing the activation of T cells in response to antigens in the perivascular space. Indeed, adventitial dendritic cells in the temporal artery remain immature in systemic inflammatory conditions that mimic giant-cell arteritis and polymyalgia rheumatica, such as fever of unknown origin.18 The inhibition of T-cell immune responses at vital tissue sites by immature dendritic cells is called immunoprivilege.37,39,40
|
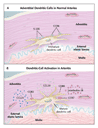
Lesions in temporal arteries from patients with giant-cell arteritis contain numerous dendritic cells in the adventitia and the inflamed wall.18,41 These dendritic cells are mature and activated (Figure 2B). They produce the inflammatory cytokines interleukin-6 and interleukin-18 and express CD86, a coreceptor required for successful interactions between T cells and dendritic cells.18 These mature dendritic cells have lost the ability to tolerize T cells; instead, they have acquired the ability to initiate and maintain the activation of T cells.
The activation of adventitial dendritic cells is an early event in the vasculitis. In temporal arteries from patients with polymyalgia rheumatica, which lack inflammatory infiltrates, the dendritic cells are no longer immature and have acquired the activation marker CD83. They also produce chemokines typically associated with fully mature dendritic cells (unpublished data).
Activated dendritic cells typically leave tissue and migrate to lymph nodes, thereby confining the activation of CD4+ T cells to lymphoid tissues.36,38,42 In polymyalgia rheumatica and giant-cell arteritis, however, activated dendritic cells remain in the artery. One mechanism that may account for this phenomenon is that dendritic cells in the inflamed artery produce the homing chemokines CCL18, CCL19, and CCL21 and express receptors for these chemokines. Large amounts of these chemokines have been identified in arteritic lesions.18 These chemokines are usually produced in lymph nodes, and they direct activated dendritic cells to secondary lymphoid tissues.42 Dendritic cells in granulomatous formations in temporal arteries in giant-cell arteritis also possess the chemokine receptor CCR7. Because CCR7 binds CCL19 and CCL21, dendritic cells are trapped in the vascular infiltrates.36,38
Granuloma Formation in Giant-Cell Arteritis
Tissue-resident T cells induce and maintain inflammatory infiltrates by
releasing interferon-![]() (Figure
3A).43,44
In giant-cell arteritis, interferon-
(Figure
3A).43,44
In giant-cell arteritis, interferon-![]() �producing
CD4+ T cells are located in the adventitia, close to resident
dendritic cells. In the media, the cellular infiltrate is organized
in granulomas, forming a unique spatial relation between activated T
cells and macrophages. The formation of granulomas is strictly
dependent on T cells and is typical of immune responses to
indigestible antigens. Studies of genetically altered animals and
patients with immunodeficiency syndromes have identified the critical
molecular components.45 Deficiency of
interferon-
�producing
CD4+ T cells are located in the adventitia, close to resident
dendritic cells. In the media, the cellular infiltrate is organized
in granulomas, forming a unique spatial relation between activated T
cells and macrophages. The formation of granulomas is strictly
dependent on T cells and is typical of immune responses to
indigestible antigens. Studies of genetically altered animals and
patients with immunodeficiency syndromes have identified the critical
molecular components.45 Deficiency of
interferon-![]() prevents the formation
of granulomas.46,47,48,49,50
Interleukin-12, a powerful inducer of interferon-
prevents the formation
of granulomas.46,47,48,49,50
Interleukin-12, a powerful inducer of interferon-![]() ,
can facilitate the formation of granulomas, but it is not essential.51,52,53
Interferon-
,
can facilitate the formation of granulomas, but it is not essential.51,52,53
Interferon-![]() is a key cytokine
in the arteritic lesion of giant-cell arteritis, and it is absent
from tissues affected by polymyalgia rheumatica, which do not have
fully developed vascular infiltrates.54 Interferon-
is a key cytokine
in the arteritic lesion of giant-cell arteritis, and it is absent
from tissues affected by polymyalgia rheumatica, which do not have
fully developed vascular infiltrates.54 Interferon-![]() mediates most of its inflammatory effects by inducing macrophages to
perform a broad spectrum of effector functions.20,55
Interestingly, interleukin-12 is missing from the vascular lesions of
giant-cell arteritis, indicating a role for alternative interferon-
mediates most of its inflammatory effects by inducing macrophages to
perform a broad spectrum of effector functions.20,55
Interestingly, interleukin-12 is missing from the vascular lesions of
giant-cell arteritis, indicating a role for alternative interferon-![]() �inducing
cytokines.18
�inducing
cytokines.18
|
The mechanisms of injury in Takayasu's arteritis are similar to those of giant-cell arteritis and include the activation of macrophages by stimulated T cells. However, in Takayasu's arteritis, T cells may have additional effector functions, such as releasing the pore-forming protein perforin, which could directly contribute to the structural damage incurred by the aortic wall.56,57
Mechanisms of Tissue Damage in Giant-Cell Arteritis
Granulomatous reactions can encapsulate and wall off immunogens, but they cause extensive tissue damage. Tissue necrosis resulting from released lytic enzymes is characteristic of Wegener's granulomatosis and Churg�Strauss vasculitis.58 Fibrinoid necrosis and destruction of the arterial wall are hallmarks of nongranulomatous vasculitides, such as panarteritis nodosa.1 Necrosis is not a feature of giant-cell arteritis, and its presence should raise doubts about the diagnosis of giant-cell arteritis.1 Thinning of the temporal-artery wall has been described, and the end stage of giant-cell aortitis may be complicated by the formation and rupture of aneurysms. However, the release of lytic enzymes is not sufficient for the overt digestion of arterial-wall components in most patients. The exception is the fragmentation of the internal elastic lamina, a process often associated with the appearance of multinucleated giant cells.
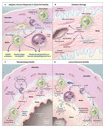
Screening for genes that are differentially expressed in inflamed temporal arteries has been fruitful in deciphering the effector mechanisms of arterial injury.59,60 Many genes that are overexpressed in inflamed arteries participate in pathways of oxidative stress. Lipid peroxidation mediated by reactive oxygen intermediates is an important pathway of arterial-wall damage (Figure 3B).59,60 The chief target of the oxidative attack is medial smooth-muscle cells. Nitrosative stress resulting from the accumulation of nitrated proteins primarily affects endothelial cells lining newly formed capillaries in the arterial media and is mediated by the concerted action of endothelial nitric oxide synthase and reactive oxygen intermediates produced by medial macrophages.61 Protein nitration can also alter intracellular signaling cascades and modulate cellular function. In healthy blood vessels, the media is free of capillaries and is inaccessible to inflammatory cells. Endothelial activation in inflammation-induced neocapillaries, brought about by oxidative stress, can increase the recruitment of lymphocytes and macrophages and aggravate inflammation.
Oxidative stress occurs in the medial layer but not in the adventitia or the intima, probably because only macrophages in the media produce reactive oxygen intermediates. The function of macrophages in the arterial wall is highly organized. Macrophages committed to restricted effector functions are confined to distinct regions,62 a finding that strongly indicates that the artery itself guides the inflammatory process. Signals derived from matrix components in microenvironments of the artery may regulate selective homing or in situ differentiation of effector macrophages.63
Oxygen-derived free radicals and their metabolites injure tissue through several mechanisms; the most important is oxidation of membrane lipids, resulting in structural disintegration and cell death. Reactive oxygen intermediates are not always cytotoxic; they can also alter cellular function by disrupting intracellular signaling cascades.64,65 Activation of gene transcription as an outcome of oxidative stress is exemplified by the induction of the keto-reductase aldose reductase in the medial layer of inflamed temporal arteries.60 In vivo, the expression of aldose reductase in medial smooth-muscle cells closely mirrors the distribution of the toxic aldehyde 4-hydroxy-2-nonenal, and 4-hydroxy-2-nonenal up-regulates the transcription of aldose reductase in vitro.60 The induction of this enzyme appears to be a protective mechanism; aldose reductase detoxifies the toxic aldehydes that are produced by oxidative stress. Blockade of aldose reductase in vivo promptly increases tissue levels of 4-hydroxy-2-nonenal and induces apoptosis of smooth-muscle cells.60 This example illustrates the balance between destructive and protective mechanisms that is characteristic of the pattern of response to injury in giant-cell arteritis. Intimal hyperplasia is favored over aneurysm formation.
Response of Arteries to Immune-Mediated Injury
Hemorrhage and the formation of aneurysms in extracranial arteries are not features of giant-cell arteritis. Instead, the most feared complications result from ischemia. Blindness is caused most often by anterior ischemic optic neuropathy.66,67 Luminal stenosis is caused by rapid, concentric hyperplasia of the intima. Rarely does thrombotic occlusion contribute to the blockage of blood flow.
Thickening of the intima is a general response of arterial vessels to injury that is shared by noninflammatory and inflammatory vasculopathies. Intimal hyperplasia, intended to heal and repair, is achieved by the mobilization and migration of myofibroblasts. The proliferation of myofibroblasts and the deposition of extracellular matrix are central events in the formation of the expanding intima (Figure 3C). Platelet-derived growth factor is abundant in the temporal arteries of patients with giant-cell arteritis.68 Interestingly, the extent of platelet-derived growth factor production in the vascular lesions correlates with the degree of luminal occlusion and the clinical manifestations of ischemia,68 such as jaw claudication and visual loss, suggesting that it is a critical growth factor for intimal myofibroblasts. Resident cells of the arterial wall � in particular, smooth-muscle cells � produce platelet-derived growth factor. This growth factor also derives from macrophages, specifically those at the media�intima junction.68
Outgrowth of the intima could not occur without accompanying angiogenesis. Medium-sized arteries receive oxygen through either diffusion from the macrolumen or the capillary network of the vasa vasorum in the adventitia. In arteries affected by giant-cell arteritis, microvessels emerge in the media and the hyperplastic intima (Figure 3C).35,69,70 The topical arrangement is strictly regulated and forms a ring-like structure in the proximal intima. The level of production of vascular endothelial growth factor but not fibroblast growth factor 2 in the vasculitic lesion correlates closely with the number of neocapillaries.68 Vascular endothelial growth factor derives from macrophages and multinucleated giant cells. Thus, a coordinated action of the arterial wall and the invading immune cells drives the hyperproliferative response of the intima.
Heterogeneity of Clinical Disease and Its Pathogenic Correlates
Pathogenic studies have shown that giant-cell arteritis is a heterogeneous
syndrome, with clinical phenotypes correlating with different
patterns of immune response. The most distinct variant is polymyalgia
rheumatica, in which vascular inflammation remains undetectable.54
In polymyalgia rheumatica, adventitial dendritic cells are mature and
both interleukin-1 and interleukin-6 are detectable, but interferon-![]() �producing
T cells are not recruited into vascular tissue. Interestingly, the
systemic inflammatory component of giant-cell arteritis and that of
polymyalgia rheumatica are indistinguishable19
and are characterized by the production of large amounts of
macrophage-derived inflammatory cytokines.71,72
�producing
T cells are not recruited into vascular tissue. Interestingly, the
systemic inflammatory component of giant-cell arteritis and that of
polymyalgia rheumatica are indistinguishable19
and are characterized by the production of large amounts of
macrophage-derived inflammatory cytokines.71,72
A clinically important subgroup of patients with medium- and large-vessel vasculitis consists of those at risk for ischemic complications.4,66,67 Arterial stenosis is closely associated with intimal hyperplasia, driven by growth and angiogenic factors.68,69 Another subgroup of patients do not have luminal stenosis � the arteritis proceeds without eliciting intimal growth (Figure 3C). 68 Mechanisms underlying the variability of the response of the artery to injury are not fully understood, but the nature of T-cell responses appears to regulate the extent of intimal hyperplasia and subsequent tissue ischemia.69,73 Morphologic hallmarks of the temporal-artery�biopsy specimens and tissue cytokine profiles could be useful in risk stratification. In patients without luminal stenosis, the diagnosis relies heavily on arterial biopsy, because imaging procedures such as ultrasonography depend on blood-flow abnormalities and lack the sensitivity to detect subtle inflammation of arterial walls.74 The diagnosis of nonstenosing giant-cell arteritis in vascular beds that are not accessible for a routine biopsy, such as the subclavian, axillary, or vertebral arteries, may require positron-emission tomography with fluorodeoxyglucose.75 This method has high sensitivity for the detection of metabolic changes in the arterial wall.76,77 Magnetic resonance imaging and computed tomography are useful, but they may not always be sensitive enough to detect arterial-wall abnormalities.78,79
Therapeutic Implications
Systemic inflammation and the arterial-wall lesions in giant-cell arteritis and polymyalgia rheumatica are intimately linked, but they may require different therapeutic approaches (Table 1). Within the two disease components, a myriad of mediators has emerged, each of which contributes to different aspects of the systemic or vascular pathogenesis. These mediators are promising targets for a tailored strategy in the treatment of medium- and large-vessel vasculitides (Table 2).
|
|
Optimizing Corticosteroid Treatment
The production of inflammatory cytokines by the innate immune system
is exquisitely sensitive to the immunosuppressive action of
corticosteroids. Inhibition of the nuclear factor-![]() B
pathway by corticosteroids disrupts a multitude of inflammatory
signals,80 which explains why symptoms
of systemic inflammation caused by stimulation of the innate immune
system are highly sensitive to corticosteroids.81,82
The immune response in the artery, however, is relatively resistant
and persists despite long-term therapy with corticosteroids.81
These two components of giant-cell arteritis may require different
therapeutic interventions (Table 1). The ability of
corticosteroids to prevent blindness83,84
may result from a reduction in vascular-wall edema or the disruption of
the triggering of dendritic cells in the arterial adventitia. Given
the limited efficacy of corticosteroids in eliminating vessel-wall
inflammation, the value of currently used regimens of corticosteroids
needs to be readdressed.
B
pathway by corticosteroids disrupts a multitude of inflammatory
signals,80 which explains why symptoms
of systemic inflammation caused by stimulation of the innate immune
system are highly sensitive to corticosteroids.81,82
The immune response in the artery, however, is relatively resistant
and persists despite long-term therapy with corticosteroids.81
These two components of giant-cell arteritis may require different
therapeutic interventions (Table 1). The ability of
corticosteroids to prevent blindness83,84
may result from a reduction in vascular-wall edema or the disruption of
the triggering of dendritic cells in the arterial adventitia. Given
the limited efficacy of corticosteroids in eliminating vessel-wall
inflammation, the value of currently used regimens of corticosteroids
needs to be readdressed.
Combination Therapy
It is unlikely that a single therapeutic intervention can abrogate giant-cell arteritis or suppress both the systemic inflammation and the vascular abnormalities. Instead, different strategies must be used for the multiple pathogenic pathways operational in the innate and adaptive immune systems. Suppressing luminal occlusion by modulating pathways of the arterial response to injury may be the next best step in improving the management of large-vessel vasculitides. Possible interventions include the prevention of oxidative injury, down-regulation of myofibroblast proliferation, and modulation of capillary formation in the arterial wall (Table 2).
Aspirin
Although thromboembolic occlusion is not a mechanism in giant-cell arteritis,
therapeutic benefit has been reported with the use of low-dose
aspirin.85 Prevention of platelet aggregation is
potentially effective, even in patients with partial luminal occlusion.
In addition, a potent antiinflammatory action of aspirin has been
described in the mouse chimera model of giant-cell arteritis. Aspirin
interferes with the transcriptional activation of the interferon-![]() gene, a mechanism of action that may be particularly useful in
inhibiting T-cell function in the granulomatous infiltrates.
gene, a mechanism of action that may be particularly useful in
inhibiting T-cell function in the granulomatous infiltrates.
Supported by grants from the National Institutes of Health
(R01 EY11916 and R01 HL63919) and by the Mayo Foundation.
Source Information
From the Departments of Medicine and Immunology, Mayo Clinic, Rochester, Minn.
References
- Bjornsson J. Histopathology of primary vasculitic disorders. In: Hoffman GS, Weyand CM, eds. Inflammatory diseases of blood vessels. New York: Marcel Dekker, 2002:255-65.
- Johnston SL, Lock RJ, Gompels MM. Takayasu arteritis: a review.
J Clin Pathol 2002;55:481-486.
[Abstract/Full Text] - Salvarani C, Cantini F, Boiardi L, Hunder GG. Polymyalgia
rheumatica and giant-cell arteritis. N Engl J Med 2002;347:261-271.
[Full Text] - Hunder GG. Epidemiology of giant-cell arteritis. Cleve Clin J Med 2002;69:Suppl 2:SII-79.
- Baldursson O, Steinsson K, Bjornsson J, Lie JT. Giant cell arteritis in Iceland: an epidemiologic and histopathologic analysis. Arthritis Rheum 1994;37:1007-1012. [ISI][Medline]
- Smetana GW, Shmerling RH. Does this patient have temporal
arteritis? JAMA 2002;287:92-101.
[Abstract/Full Text] - Ghanchi FD, Dutton GN. Current concepts in giant cell (temporal) arteritis. Surv Ophthalmol 1997;42:99-123. [CrossRef][ISI][Medline]
- Brack A, Martinez-Taboada V, Stanson A, Goronzy JJ, Weyand CM. Disease pattern in cranial and large-vessel giant cell arteritis. Arthritis Rheum 1999;42:311-317. [CrossRef][ISI][Medline]
- Gravanis MB. Giant cell arteritis and Takayasu aortitis: morphologic, pathogenetic and etiologic factors. Int J Cardiol 2000;75:Suppl 1:S21-S33. [ISI][Medline]
- Evans JM, Bowles CA, Bjornsson J, Mullany CJ, Hunder GG. Thoracic aortic aneurysm and rupture in giant cell arteritis: a descriptive study of 41 cases. Arthritis Rheum 1994;37:1539-1547. [Erratum, Arthritis Rheum 1995;38:290.] [ISI][Medline]
- Weyand CM, Hunder NN, Hicok KC, Hunder GG, Goronzy JJ. HLA-DRB1 alleles in polymyalgia rheumatica, giant cell arteritis, and rheumatoid arthritis. Arthritis Rheum 1994;37:514-520. [ISI][Medline]
- Roche NE, Fulbright JW, Wagner AD, Hunder GG, Goronzy JJ, Weyand CM. Correlation of interleukin-6 production and disease activity in polymyalgia rheumatica and giant cell arteritis. Arthritis Rheum 1993;36:1286-1294. [ISI][Medline]
- Hall S, Barr W, Lie JT, Stanson AW, Kazmier FJ, Hunder GG. Takayasu arteritis: a study of 32 North American patients. Medicine (Baltimore) 1985;64:89-99. [ISI][Medline]
- Numano F, Okawara M, Inomata H, Kobayashi Y. Takayasu's arteritis. Lancet 2000;356:1023-1025. [CrossRef][ISI][Medline]
- Kerr GS. Takayasu's arteritis. Rheum Dis Clin North Am 1995;21:1041-1058. [ISI][Medline]
- Weyand CM, Goronzy JJ. Giant cell arteritis: pathogenesis. In: Hoffman GS, Weyand CM, eds. Inflammatory diseases of blood vessels. New York: Marcel Dekker, 2002:413-23.
- Weyand CM, Goronzy JJ. Pathogenic mechanisms in giant cell arteritis. Cleve Clin J Med 2002;69:Suppl 2:SII-28.
- Krupa WM, Dewan M, Jeon M-S, et al. Trapping of misdirected
dendritic cells in the granulomatous lesions of giant cell arteritis. Am J
Pathol 2002;161:1815-1823.
[Abstract/Full Text] - Wagner AD, Goronzy JJ, Weyand CM. Functional profile of tissue-infiltrating and circulating CD68+ cells in giant cell arteritis: evidence for two components of the disease. J Clin Invest 1994;94:1134-1140. [ISI][Medline]
- Weyand CM, Goronzy JJ. Arterial wall injury in giant cell arteritis. Arthritis Rheum 1999;42:844-853. [CrossRef][ISI][Medline]
- Weyand CM, Goronzy JJ. Giant cell arteritis as an antigen-driven disease. Rheum Dis Clin North Am 1995;21:1027-1039. [ISI][Medline]
- Brack A, Geisler A, Martinez-Taboada VM, Younge BR, Goronzy JJ, Weyand CM. Giant cell vasculitis is a T cell-dependent disease. Mol Med 1997;3:530-543. [ISI][Medline]
- Grunewald J, Andersson R, Rydberg L, et al. CD4+ and CD8+ T cell expansions using selected TCR V and J gene segments at the onset of giant cell arteritis. Arthritis Rheum 1994;37:1221-1227. [ISI][Medline]
- Weyand CM, Hicok KC, Hunder GG, Goronzy JJ. The HLA-DRB1 locus as a genetic component in giant cell arteritis: mapping of a disease-linked sequence motif to the antigen binding site of the HLA-DR molecule. J Clin Invest 1992;90:2355-2361. [ISI][Medline]
- Weyand CM, Schonberger J, Oppitz U, Hunder NN, Hicok KC, Goronzy JJ. Distinct vascular lesions in giant cell arteritis share identical T cell clonotypes. J Exp Med 1994;179:951-960. [Abstract]
- Martinez-Taboada V, Hunder NN, Hunder GG, Weyand CM, Goronzy JJ. Recognition of tissue residing antigen by T cells in vasculitic lesions of giant cell arteritis. J Mol Med 1996;74:695-703. [CrossRef][ISI][Medline]
- Elling P, Olsson AT, Elling H. Synchronous variations of the incidence of temporal arteritis and polymyalgia rheumatica in different regions of Denmark: association with epidemics of Mycoplasma pneumoniae infection. J Rheumatol 1996;23:112-119. [ISI][Medline]
- Elling H, Olsson AT, Elling P. Human Parvovirus and giant cell arteritis: a selective arteritic impact? Clin Exp Rheumatol 2000;18:Suppl 20:S12-S14. [Medline]
- Duhaut P, Bosshard S, Calvet A, et al. Giant cell arteritis, polymyalgia rheumatica, and viral hypotheses: a multicenter, prospective case-control study. J Rheumatol 1999;26:361-369. [ISI][Medline]
- Mitchell BM, Font RL. Detection of varicella zoster virus DNA
in some patients with giant cell arteritis. Invest Ophthalmol Vis Sci
2001;42:2572-2577.
[Abstract/Full Text] - Helweg-Larsen J, Tarp B, Obel N, Baslund B. No evidence of
parvovirus B19, Chlamydia pneumoniae or human herpes virus infection in
temporal artery biopsies in patients with giant cell arteritis. Rheumatology
(Oxford) 2002;41:445-449.
[Abstract/Full Text] - Regan MJ, Wood BJ, Hsieh YH, et al. Temporal arteritis and Chlamydia pneumoniae: failure to detect the organism by polymerase chain reaction in ninety cases and ninety controls. Arthritis Rheum 2002;46:1056-1060. [CrossRef][ISI][Medline]
- Wawryk SO, Ayberk H, Boyd AW, Rode J. Analysis of adhesion molecules in the immunopathogenesis of giant cell arteritis. J Clin Pathol 1991;44:497-501. [Abstract]
- Lavignac C, Jauberteau-Marchan MO, Liozon E, Vidal E, Catanzano G, Liozon F. Etude immunohistochimique des l�sions d'art�rite temporale de Horton avant et au cours de la corticoth�rapie. Rev Med Interne 1996;17:814-820. [ISI][Medline]
- Cid MC, Cebrian M, Font C, et al. Cell adhesion molecules in the development of inflammatory infiltrates in giant cell arteritis: inflammation-induced angiogenesis as the preferential site of leukocyte-endothelial cell interactions. Arthritis Rheum 2000;43:184-194. [CrossRef][ISI][Medline]
- Caux C, Vanbervliet B, Massacrier C, et al. Regulation of dendritic cell recruitment by chemokines. Transplantation 2002;73:Suppl:S7-S11. [ISI][Medline]
- Huang FP, MacPherson GG. Continuing education of the immune system � dendritic cells, immune regulation and tolerance. Curr Mol Med 2001;1:457-68.
- Randolph GJ. Dendritic cell migration to lymph nodes: cytokines, chemokines, and lipid mediators. Semin Immunol 2001;13:267-274. [CrossRef][ISI][Medline]
- Steinman RM, Nussenzweig MC. Avoiding horror autotoxicus: the
importance of dendritic cells in peripheral T cell tolerance. Proc Natl Acad
Sci U S A 2002;99:351-358.
[Abstract/Full Text] - Guermonprez P, Valladeau J, Zitvogel L, Thery C, Amigorena S. Antigen presentation and T cell stimulation by dendritic cells. Annu Rev Immunol 2002;20:621-667. [CrossRef][ISI][Medline]
- Cid MC, Campo E, Ercilla G, et al. Immunohistochemical analysis of lymphoid and macrophage cell subsets and their immunologic activation markers in temporal arteritis: influence of corticosteroid treatment. Arthritis Rheum 1989;32:884-893. [ISI][Medline]
- Cyster JG. Leukocyte migration: scent of the T zone. Curr Biol 2000;10:R30-R33. [CrossRef][ISI][Medline]
- Wagner AD, Bjornsson J, Bartley GB, Goronzy JJ, Weyand CM. Interferon-gamma-producing T cells in giant cell vasculitis represent a minority of tissue-infiltrating cells and are located distant from the site of pathology. Am J Pathol 1996;148:1925-1933. [Abstract]
- Blain H, Abdelmouttaleb I, Belmin J, et al. Arterial wall
production of cytokines in giant cell arteritis: results of a pilot study
using human temporal artery cultures. J Gerontol A Biol Sci Med Sci
2002;57:M241-M245.
[Abstract/Full Text] - Lammas DA, De Heer E, Edgar JD, et al. Heterogeneity in the granulomatous response to mycobacterial infection in patients with defined genetic mutations in the interleukin 12-dependent interferon-gamma production pathway. Int J Exp Pathol 2002;83:1-20. [CrossRef][ISI][Medline]
- Cooper AM, Dalton DK, Stewart TA, Griffin JP, Russell DG, Orme IM. Disseminated tuberculosis in interferon gamma gene-disrupted mice. J Exp Med 1993;178:2243-2247. [Abstract]
- Flynn JL, Chan J, Triebold KJ, Dalton DK, Stewart TA, Bloom BR. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J Exp Med 1993;178:2249-2254. [Abstract]
- Kamijo R, Le J, Shapiro D, et al. Mice that lack the interferon-gamma receptor have profoundly altered responses to infection with Bacillus Calmette-Guerin and subsequent challenge with lipopolysaccharide. J Exp Med 1993;178:1435-1440. [Abstract]
- Dorman SE, Holland SM. Mutation in the signal-transducing chain
of the interferon-gamma receptor and susceptibility to mycobacterial
infection. J Clin Invest 1998;101:2364-2369.
[Abstract/Full Text] - Jouanguy E, Altare F, Lamhamedi S, et al. Interferon-
 -receptor
deficiency in an infant with fatal bacille Calmette-Gu�rin infection. N
Engl J Med 1996;335:1956-1961.
-receptor
deficiency in an infant with fatal bacille Calmette-Gu�rin infection. N
Engl J Med 1996;335:1956-1961.
[Full Text] - Altare F, Lammas D, Revy P, et al. Inherited interleukin 12
deficiency in a child with bacille Calmette-Guerin and Salmonella
enteritidis disseminated infection. J Clin Invest 1998;102:2035-2040.
[Abstract/Full Text] - de Jong R, Altare F, Haagen IA, et al. Severe mycobacterial and
Salmonella infections in interleukin-12 receptor-deficient patients. Science
1998;280:1435-1438.
[Abstract/Full Text] - Altare F, Durandy A, Lammas D, et al. Impairment of
mycobacterial immunity in human interleukin-12 receptor deficiency. Science
1998;280:1432-1435.
[Abstract/Full Text] - Weyand CM, Hicok KC, Hunder GG, Goronzy JJ. Tissue cytokine patterns in patients with polymyalgia rheumatica and giant cell arteritis. Ann Intern Med 1994;121:484-491. [ISI][Medline]
- Weyand CM. The Dunlop-Dottridge Lecture: the pathogenesis of giant cell arteritis. J Rheumatol 2000;27:517-522. [ISI][Medline]
- Seko Y, Sato O, Takagi A, et al. Restricted usage of T-cell
receptor Valpha-Vbeta genes in infiltrating cells in aortic tissue of
patients with Takayasu's arteritis. Circulation 1996;93:1788-1790.
[Abstract/Full Text] - Seko Y, Minota S, Kawasaki A, et al. Perforin-secreting killer cell infiltration and expression of a 65-kD heat-shock protein in aortic tissue of patients with Takayasu's arteritis. J Clin Invest 1994;93:750-758. [ISI][Medline]
- Sneller MC. Granuloma formation, implications for the pathogenesis of vasculitis. Cleve Clin J Med 2002;69:Suppl 2:SII-40.
- Rittner HL, Kaiser M, Brack A, Szweda LI, Goronzy JJ, Weyand
CM. Tissue-destructive macrophages in giant cell arteritis. Circ Res
1999;84:1050-1058.
[Abstract/Full Text] - Rittner HL, Hafner V, Klimiuk PA, Szweda LI, Goronzy JJ, Weyand
CM. Aldose reductase functions as a detoxification system for lipid
peroxidation products in vasculitis. J Clin Invest 1999;103:1007-1013.
[Abstract/Full Text] - Borkowski A, Younge BR, Szweda L, et al. Reactive nitrogen
intermediates in giant cell arteritis: selective nitration of neocapillaries.
Am J Pathol 2002;161:115-123.
[Abstract/Full Text] - Weyand CM, Wagner AD, Bjornsson J, Goronzy JJ. Correlation of
the topographical arrangement and the functional pattern of
tissue-infiltrating macrophages in giant cell arteritis. J Clin Invest
1996;98:1642-1649.
[Abstract/Full Text] - Weyand CM. Vasculitis: a dialogue between the artery and the immune system. In: Hoffman GS, Weyand CM, eds. Inflammatory diseases of blood vessels. New York: Marcel Dekker, 2002:1-12.
- Goldschmidt-Clermont PJ, Moldovan L. Stress, superoxide, and signal transduction. Gene Expr 1999;7:255-260. [ISI][Medline]
- Droge W. Free radicals in the physiological control of cell
function. Physiol Rev 2002;82:47-95.
[Abstract/Full Text] - Hayreh SS. Anterior ischemic optic neuropathy. Clin Neurosci 1997;4:251-263. [ISI][Medline]
- McFadzean RM. Ischemic optic neuropathy and giant cell arteritis. Curr Opin Ophthalmol 1998;9:10-17. [Medline]
- Kaiser M, Weyand CM, Bjornsson J, Goronzy JJ. Platelet-derived growth factor, intimal hyperplasia, and ischemic complications in giant cell arteritis. Arthritis Rheum 1998;41:623-633. [CrossRef][ISI][Medline]
- Kaiser M, Younge B, Bjornsson J, Goronzy JJ, Weyand CM.
Formation of new vasa vasorum in vasculitis: production of angiogenic
cytokines by multinucleated giant cells. Am J Pathol 1999;155:765-774.
[Abstract/Full Text] - Nelimarkka L, Salminen H, Kuopio T, et al. Decorin is produced
by capillary endothelial cells in inflammation-associated angiogenesis. Am J
Pathol 2001;158:345-353.
[Abstract/Full Text] - Weyand CM, Fulbright JW, Evans JM, Hunder GG, Goronzy JJ.
Corticosteroid requirements in polymyalgia rheumatica. Arch Intern Med
1999;159:577-584.
[Abstract/Full Text] - Weyand CM, Fulbright JW, Hunder GG, Evans JM, Goronzy JJ. Treatment of giant cell arteritis: interleukin-6 as a biologic marker of disease activity. Arthritis Rheum 2000;43:1041-1048. [CrossRef][ISI][Medline]
- Weyand CM, Tetzlaff N, Bjornsson J, Brack A, Younge B, Goronzy JJ. Disease patterns and tissue cytokine profiles in giant cell arteritis. Arthritis Rheum 1997;40:19-26. [ISI][Medline]
- Schmidt WA. Doppler ultrasonography in the diagnosis of giant cell arteritis. Clin Exp Rheumatol 2000;18:Suppl 20:S40-S42. [ISI][Medline]
- Blockmans D, Maes A, Stroobants S, et al. New arguments for a vasculitic nature of polymyalgia rheumatica using positron emission tomography. Rheumatology (Oxford) 1999;38:444-447. [Abstract]
- Blockmans D, Stroobants S, Maes A, Mortelmans L. Positron emission tomography in giant cell arteritis and polymyalgia rheumatica: evidence for inflammation of the aortic arch. Am J Med 2000;108:246-249. [CrossRef][ISI][Medline]
- Turlakow A, Yeung HW, Pui J, et al. Fludeoxyglucose positron
emission tomography in the diagnosis of giant cell arteritis. Arch Intern
Med 2001;161:1003-1007.
[Abstract/Full Text] - Atalay MK, Bluemke DA. Magnetic resonance imaging of large vessel vasculitis. Curr Opin Rheumatol 2001;13:41-47. [CrossRef][ISI][Medline]
- Stanson AW. Imaging findings in extracranial (giant cell) temporal arteritis. Clin Exp Rheumatol 2000;18:Suppl 20:S43-S48. [ISI][Medline]
- Yamamoto Y, Gaynor RB. Therapeutic potential of inhibition of
the NF-kappaB pathway in the treatment of inflammation and cancer. J Clin
Invest 2001;107:135-142.
[Full Text] - Brack A, Rittner HL, Younge BR, Kaltschmidt C, Weyand CM,
Goronzy JJ. Glucocorticoid-mediated repression of cytokine gene
transcription in human arteritis-SCID chimeras. J Clin Invest
1997;99:2842-2850.
[Abstract/Full Text] - Weyand CM, Kaiser M, Yang H, Younge B, Goronzy JJ. Therapeutic effects of acetylsalicylic acid in giant cell arteritis. Arthritis Rheum 2002;46:457-466. [CrossRef][ISI][Medline]
- Achkar AA, Lie JT, Hunder GG, O'Fallon WM, Gabriel SE. How does previous corticosteroid treatment affect the biopsy findings in giant cell (temporal) arteritis? Ann Intern Med 1994;120:987-992. [ISI][Medline]
- Ray-Chaudhuri N, Kine DA, Tijani SO, et al. Effect of prior
steroid treatment on temporal artery biopsy findings in giant cell arteritis.
Br J Ophthalmol 2002;86:530-532.
[Abstract/Full Text] - Liozon E, Herrmann F, Ly K, et al. Risk factors for visual loss in giant cell (temporal) arteritis: a prospective study of 174 patients. Am J Med 2001;111:211-217. [CrossRef][ISI][Medline]